Table
des matières - ![]() Précédente
- Suivante
Précédente
- Suivante![]()
T. nigrescens a pu se reproduire sur quelques-unes des espèces d'animaux hôtes qui lui ont été proposées. Dans la mesure où l'on peut exclure l'hypothèse d'une reproduction du prédateur sur un substrat végétal en l'absence d'animaux hôtes (5.4), T. nigrescens a nécessairement dévoré un certain nombre d'individus des animaux hôtes pour assurer sa descendance. En règle générale, on a effectivement trouvé dans les préparations expérimentales contenant le prédateur un nombre d'animaux hôtes inférieur à ce qu'il était dans les préparations témoins sans T. nigrescens. D'un point de vue statistique toutefois, celui-ci n'a pu inhiber l'accroissement des populations d'animaux hôtes que sur un petit nombre de monocultures. Au cours des séries d'essais faisant intervenir, sur du mais, la souche de laboratoire de L. oryzae d'une part, et R. dominica d'autre part, on a même compté en moyenne dans les préparations contenant le prédateur un nombre d'animaux hôtes siginificativement plus élevé que dans les préparations témoins.
On serait tenté de croire que pour pouvoir exercer une influence inhibitrice significative sur la population d'animaux hôtes, il faudrait impérativement que de nombreux individus de la génération F1 du prédateur se développent dans les préparations. Selon REES (1985), une larve de T. nigrescens détruit à peu près 60 larves de P. truncatus pour assurer son évolution jusqu'au stade de l'imago. Les quelques exemples qui suivent prouvent cependant que cette seule explication est insuffisante.
Bien que l'on ait dénombré dans les élevages de R. dominica sur du mars (tabl. 3.1. et 3.2) plus de descendants de T. nigrescens que dans ceux de P. truncatus placés sur du manioc, du sorgho et du blé (tabl. 1), on n'a pas pu mettre en évidence d'effets inhibiteurs sur l'accroissement des populations de R. dominica, alors que les populations de P. truncatus placées sur les substrats cités avaient été significativement influencées par T. nigrescens.
Dans les élevages de O. surinamensis sur les flocons d'avoine, le manioc et le maïs, on n'a compté là encore qu'un petit nombre d'individus F1 du prédateur, bien que l'on ait observé des effets significatifs de la part de T. nigrescens.
Un calcul arithmétique va nous permettre d'expliquer la forte influence exercée par T. nigrescens sur la population d'animaux hôtes, alors même que l'on n'y a trouvé qu'une descendance réduite du prédateur, voire pas de descendance du tout. Selon LELIVELDT (1990), un imago de T. nigrescens dévore en moyenne 5,7 oeufs ou 4,9 larves de P. truncatus par jour. REES (1985) a déterminé quant à lui une moyenne de 1,7 larves de P. truncatus Etant donné que les 10 imagos de T. nigrescens utilisés étaient en général encore vivants à l'issue des 2 mois d'essai (60 jours), les imagos seraient, selon LELIVELDT (1990), capables de dévorer à eux seuls 10 x 5,7 x 60 (=3420) oeufs et 10 x 4,9 x 60 (=2940) larves de P. truncatus. Selon REES (1985), on arriverait ici à un nombre de 10 x 1,7 x 60 (=1020) larves de P. truncatus au cours de la période d'essai.
Dû à leur haute longévité, les imagos de T. nigrescens seraient par conséquent en mesure de contribuer très fortement à la décimation des animaux hôtes, même si, selon REES (1985), ils ne dévorent en 24 h que la moitié des proies détruites par les larves du prédateur.
Nous indiquerons ici à titre d'exemples deux cas concrets qui illustrent ces calculs arithmétiques. En dépit du fait que T. nigrescens ne s'est pas reproduit au cours des séries d'essais avec 0. mercator (tabl. 3.7) et T. granarium sur du son (tabl. 3.9), on trouvé dans les échantillons contenant le prédateur un nombre d'animaux hôtes significativement inférieur à celui des préparations témoins. Il faut donc admettre que les imagos de T. nigrescens placés à l'origine dans les préparations ont dévoré les individus de 0. mercator et de T. granarium.
Au contraire de leurs larves, les T. nigrescens adultes ne sont pas dépendants d'une alimentation animale, mais peuvent se nourrir de substrats végétaux riches en amidon. Lorsque les imagos du prédateur placés dans les élevages d'animaux hôtes accordent la préférence à une nourriture végétale au détriment des proies animales proposées, leur influence sur l'évolution des populations d'animaux hôtes sera moindre que si les T. nigrescens des générations P et F1 chassent ensemble.
C'est ce phénomène qui explique que l'on n'ait pas pu observer d'influence visible du prédateur sur la population d'animaux hôtes, bien que l'on ait constaté la présence dans les échantillons d'une descendance relativement nombreuse de T. nigrescens. Les deux séries d'essais mettant en oeuvre R. dominica sur du mais (tabl. 3. I et 3.2) nous en fournissent des exemples.
Il est assez improbable que la population de R. dominica ait été en mesure de compenser les pertes causées par T. nigrescens par un taux de ponte renforcé.
Il est certain que la biologie, de même que la morphologie et le comportement des organismes destinés eux-mêmes à servir de proies jouent un rôle important lorsqu'il s'agit de déterminer si T. nigrescens est en mesure d'exploiter pour sa nutrition telle ou telle espèce de coléoptères proposée comme animal hôte.
Les larves de certaines espèces de coléoptères sont très mobiles, ce qui leur permet de se soustraire aisément à l'attaque du prédateur. Ceci pourrait expliquer que les larves des Ténébrionidés et celles de certains Cucujidés aient été épargnées par T. nigrescens. Certaines larves à la mobilité assez réduite, comme celles de P. truncatus et celles d'autres bostryches, ainsi que les larves du Mycétophagidé T. stercorea, représentent au contraire pour T. nigrescens des proies faciles.
D'autres larves sont bien protégées contre les prédateurs grâce à leur cuticule sclérifiée, ou encore par leurs poils ou brosses. Les larves de P. truncatus D. porcellus, R. dominica ou T. stercorea sont en revanche glabres et relativement molles, et donc plus vulnérables que les larves de A. diaperinus, par ex., ou celles des deux espèces Tribolium, ou encore les larves de Dermestidés, qui présentent une forte pilosité.
La larve de A. diaperinus atteint en outre une taille qui exclut pour T. nigrescens toute possibilité d'en faire sa proie. Les larves d'espèces de coléoptères plus petites, ainsi par ex. celles de 0. surinamensis et de 0. mercator, peuvent au contraire représenter, même au stade prénymphal, c'est-à-dire au maximum de leur croissance, des proies acceptables pour T. nigrescens.
Parmi les larves d'animaux hôtes proposées, certaines possèdent la faculté de se défendre activement contre l'ennemi qui les menace, voire celle de contre-attaquer elles-mêmes.
La plupart des larves des coléoptères ravageurs des stocks sont dotées de mandibules puissantes, qu'elles ne réservent probablement pas au seul broyage des aliments, mais qui leur servent également à se défendre contre un ennemi cherchant à les dévorer.
A l'instar de nombreuses autres espèces de Dermestidés, les larves du trogoderme du grain (T. granarium) possèdent des épines constituant une arme efficace en cas de menace (MA et al., 1978).
Nombre d'espèces de Ténébrionidés dégagent une forte odeur. Il pourrait s'agir en l'occurrence de sécrétions d'avertissement ou de défense (allomones), que T. nigrescens est en mesure de percevoir comme telles. Ces proies perdent alors pour le prédateur leur caractère attrayant et sont épargnées.
Il est par ailleurs possible que l'ennemi évite à l'avenir de consommer des animaux de la même espèce dans la mesure où il leur a trouvé à l'origine un goût désagréable. Un tel phénomène a d'ailleurs été observé chez les punaises, qui se protègent ainsi de certains prédateurs comme les oiseaux, les reptiles et les batraciens.
Le mode biologique d'incubation des animaux hôtes a également des répercussions sur la capacité reproductive de T. nigrescens sur les diverses espèces.
Les espèces Sitophilus déposent en général leurs oeufs un par un, dans une galerie longiligne qu'elles creusent elles-même à l'intérieur des grains de substrat. L'entrée de cette galerie de ponte est hermétiquement obstruée par une goutte de sécrétion transparente (HElNZE, 1983). Du fait que T. nigrescens a beaucoup de difficulté à ronger les grains de substrat (5.3), les coléoptères peuvent se développer en toute quiétude dans l'enveloppe dure de la graine, à l'abri des prédateurs. Il est par conséquent probable que T. nigrescens ne pourrait pas davantage menacer les larves des Bruchidés.
P. truncatus, en revanche, dépose ses oeufs par groupes, dans l'une des galeries de forage qu'il creuse dans le grain de substrat, sans refermer la voie menant à sa ponte. Ces galeries de forage ont un diamètre largement suffisant pour permettre à T. nigrescens en quête de proies d'atteindre très facilement tout d'abord les oeufs et, ultérieurement, les larves.
Les galeries de forage des deux autres espèces de Bostrychidés mises en oeuvre (D. porcellus et R. dominica) étant plus petites, on peut imaginer qu'elles sont inaccessibles, ou du moins difficilement accessibles, aux imagos de T. nigrescens.
En l'absence d'une quantité suffisante de proies, les femelles de T. nigrescens ne pondent probablement qu'un nombre d'oeufs réduit. C'est ce qui explique que l'on ne trouve que peu de larves du prédateur au sein des populations, alors que le diamètre de leur corps et leur silhouette longiligne leur permettent de se glisser aisément dans les galeries de forage pour parvenir à la ponte. La ponte des espèces de coléoptères de taille réduite est par conséquent plus ou moins à l'abri du prédateur.
Outre les larves prénymphales (L2) et les imagos F1 de T. nigrescens, on a uniquement trouvé dans les échantillons des oeufs, des larves primaires ou des nymphes du prédateur. Ces résultats pourraient s'expliquer par les comportements suivants, chez les animaux hôtes ou chez le prédateur. Certaines des espèces d'animaux hôtes proposées sont des prédateurs facultatifs. En cas de pénurie de nourriture ou de surpopulation, T. castaneum, par ex., ne dévore pas seulement les individus de sa propre espèce, mais va également s'attaquer aux oeufs et aux nymphes d'autres espèces (HEINZE, 1983). Il se pourrait ainsi qu'après l'éclosion de la génération F1 les oeufs et nymphes de T. nigrescens aient été détruits par les animaux hôtes.
Autre hypothèse: les oeufs de T. nigrescens pondus dès le début des essais ont donné naissance à des larves primaires, lesquelles ont pu se développer sans être dérangées et atteindre ainsi le stade L2. La tension engendrée au sein de la population du prédateur par la situation de concurrence alimentaire qui s'en est suivie dans les conteneurs d'essais, qui étaient relativement étroits, se serait aggravée au point de conduire au cannibalisme. Il est donc possible que les oeufs pondus ultérieurement, de même que les larves primaires et les nymphes sans défense, aient été victimes de leurs propres congénères.
Les larves de T. nigrescens déjà parvenues à maturité n'ont pas trouvé dans les bocaux d'essais surpeuplés d'endroit adéquat pour effectuer leur nymphose en toute quiétude, ce qui explique que l'on ait surtout trouvé dans de nombreux échantillons des larves L2 du prédateur.
Dans le cadre de la série d'essais qui mettait en oeuvre des ravageurs primaires sur du maïs, on a trouvé dans les échantillons contenant P. truncatus davantage d'imagos F1 que de L2 de T. nigrescens.
Selon LELIVELDT (1990) et REES (1985), les larves de T. nigrescens se dissimulent pour la nymphose à l'intérieur de grains de maïs évidés. Apparemment, seul P. truncatus avait pu évider les grains de mais avec une telle efficacité au cours des 8 semaines qu'a duré l'essai, pour permettre aux larves de trouver une cachette idéale et d'y effectuer leur nymphose. Les cavités alimentaires et cavités d'incubation de R. dominica, D. porcellus, de même que celles des espèces Sitophilus étaient visiblement trop petites, ou pour quelque autre raison inappropriées, pour offrir aux larves nymphales de T. nigrescens un espace suffisant. L'hypothèse selon laquelle les grains de sorgho ou de blé rendraient difficile, voire impossible, la nymphose des larves du prédateur, et cela en raison de leur taille réduite, a été étayée par les résultats de plusieurs séries d'essais.
Aussi bien dans les élevages de P. truncatus sur du sorgho qu'au sein des populations de S. oryzae placées sur du blé, on a trouvé essentiellement des L2 du prédateur. Dans les élevages de O. surinamensis sur des flocons d'avoine, il s'est nettement avéré que les larves du prédateur avaient également de grosses difficultés à trouver un endroit adéquat pour effectuer leur nymphose.
Dans les échantillons comportant des ravageurs secondaires, on avait ajouté aux grains de maïs entiers de la mouture. Les larves de T. nigrescens sont en effet capables d'utiliser la mouture de mais pour confectionner des chambres nymphales. Cette même faculté pourrait être à l'origine du nombre relativement élevé d'imagos F1 prédateur trouvés dans les préparations contenant O. mercator, O. surinamensis et T. stercorea.
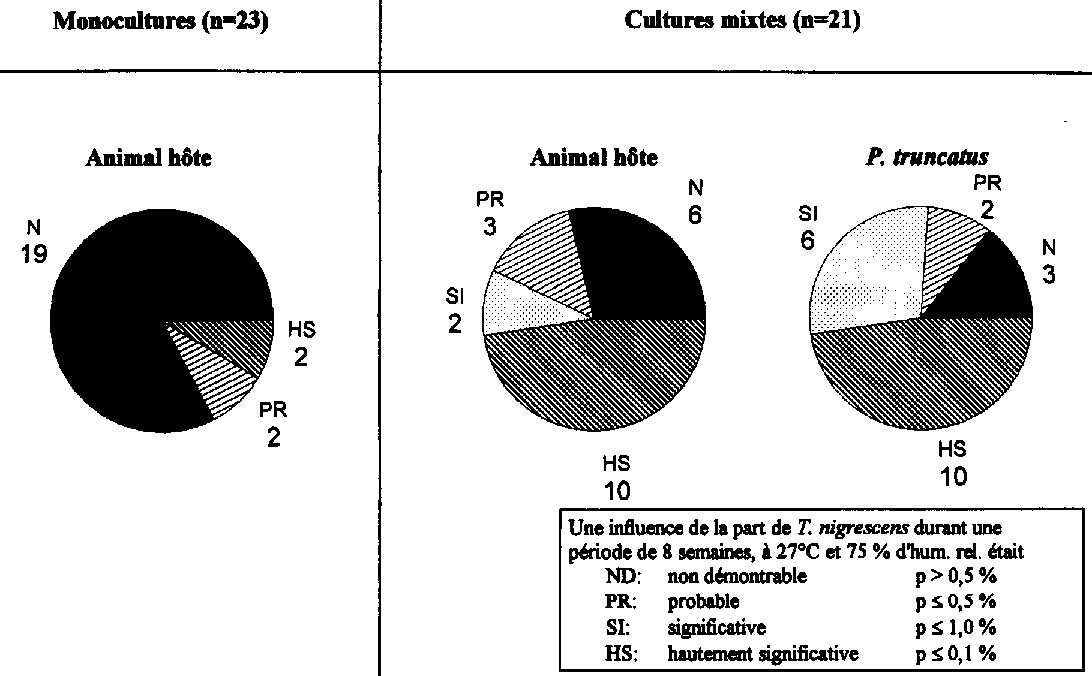
Ces résultats montrent que T. nigrescens accepte plus facilement les espèces de coléoptères offertes comme proies lorsque P. truncatus est présent dans les préparations que dans les monocultures. C'est ainsi que la plupart des espèces d'animaux hôtes placées dans des monocultures sur du mais n'ont pas été influencées par T. nigrescens, alors que dans les cultures mixtes, le nombre d'individus de ces espèces avait été réduit de façon significative par le prédateur (fig. 14), phénomène également observé par REES (1987) et LELIVELDT (1990)
Fig. 14: Influence de T. nigrescens sur le développement de populations d'animaux hôtes sur du maïs
Selon BÖYE (1988), ces résultats pourraient être dus à l'enchaînement de faits suivant. Grâce à la phéromone d'agrégation émise par P. truncatus T. nigrescens est en mesure de sentir sa présence à une grande distance et de le rechercher activement (REES et al., 1990; REES, 1990). Il est probable que la phéromone émise par le bostrychidé agit sur T. nigrescens comme un "allelo-chemical", stimulant ainsi chez le prédateur un comportement de recherche et de consommation de la proie au sein de cultures mixtes. Espérant rencontrer au cours de sa quête de nourriture sa proie favorite, P. truncatus T. nigrescens dépiste et dévore également les oeufs et les larves d'autres animaux hôtes se trouvant dans le conteneur d'essais.
Dans la plupart des cas, cependant, on n'est pas parvenu à déterminer avec certitude si T. nigrescens avait vraiment utilisé comme proies les individus des espèces de coléoptères présentes dans les cultures mixtes ou s'il s'était contenté de les tuer. Les résultats de deux séries d'essais indiquent toutefois que T. nigrescens a effectivement dévoré les animaux hôtes.
Dans les cultures mixtes comportant des T. castaneum et des T. confusum provenant du Togo (tabl. 3.5 et 3.6), aucune influence sur la population de P. truncatus n'a été observée, T. nigrescens ayant au contraire réduit de manière significative le nombre des imagos de Tribolium. Dans le premier cas, P. truncatus n'a pas pu se reproduire du tout, dans le second, la reproduction est demeurée insignifiante. Neuf à dix descendants de T. nigrescens ont été dénombrés en moyenne dans ces préparations. Etant donné que les individus de P. truncatus étaient totalement absents de ces préparations ou ne s'y trouvaient qu'en petit nombre, on peut supposer que le prédateur a utilisé comme source de nourriture la seconde espèce d'animaux hôtes et qu'il s'est également reproduit sur elle.
T. nigrescens s'est mieux reproduit dans la plupart des cultures mixtes qu'au sein des populations de P. truncatus d'importance comparable. REES (1987) et LELIVELDT (1990) ont eux aussi observé chez le prédateur une capacité reproductive renforcée sur des cultures mixtes de ce type. Il se pourrait que le phénomène de concurrence pousse les animaux hôtes à pondre davantage d'oeufs afin d'assurer la survie de leur propre espèce. T. nigrescens aurait eu ainsi davantage de proies adéquates à sa disposition que par exemple dans les monocultures de P. truncatus
Il est également plausible que l'ingestion de proies appartenant à diverses espèces ait contribué à une amélioration qualitative de l'alimentation du prédateur, ce qui se serait traduit par un taux de multiplication renforcé.
Selon LELIVELDT (1990), T. nigrescens s'attaque aussi en période de disette à des imagos P. truncatus fraîchement éclos. Il a été établi dans deux cas que T. nigrescens avait également dévoré des imagos d'autres espèces de coléoptères.
R. dominica n'a pas été en mesure d'exploiter le manioc comme substrat d'incubation. En comparaison des préparations témoins, on a dénombré dans les échantillons contenant T. nigrescens une réduction hautement significative du nombre des animaux hôtes adultes (tabl. 3.1). Ceci permet de conclure que le prédateur a également accepté comme proies des imagos adultes de R dominica. Vu que tous les animaux ont été congelés avant l'évaluation, il n'est plus possible de dire si T. nigrescens a tué les R. dominica adultes ou s'il les a dévorés après leur mort seulement, ce qui fait que l'on ignore combien, parmi les imagos de R. dominica dénombrés à la fin de l'essai dans les témoins, étaient déjà morts auparavant.
Il est improbable que ce phénomène puisse encore s'expliquer par une réaction de cannibalisme chez les imagos de R. dominica, réaction provoquée par la présence de T. nigrescens.
Un cas analogue a également été observé dans les cultures mixtes sur lesquelles on avait placé P. ratzeburgi (tabl. 3.5). Les imagos de P. ratzeburgi ne se sont pas reproduits dans les préparations témoins. Le nombre d'animaux hôtes adultes de la génération P trouvés dans les échantillons comportant le prédateur avait diminué de manière hautement significative. Dans les populations mixtes composées de P. ratzeburgi et de P. truncatus T. nigrescens a par conséquent accepté les adultes de P. ratzeburgi comme proies.
Dans les 6 autres séries d'essais entreprises (avec D. porcellus et O. surinamensis sur du manioc, ainsi que sur des cultures mixtes réunissant D. porcellus, O. mercator, C. dimidiatus ou T. granarium sur du maïs), on a compté dans les préparations expérimentales un nombre d'animaux hôtes adultes inférieur à celui qui avait été introduit à l'origine. Dans tous les cas évoqués ici, on n'a observé dans la plupart des préparations témoins qu'une faible activité de reproduction de la part des animaux hôtes.
Si l'on peut en déduire selon toute probabilité que T. nigrescens a accepté dans les cultures mixtes les imagos de la totalité de ces espèces d'animaux hôtes comme proies, cette conclusion n'est toutefois pas démontrable. Parmi les animaux hôtes adultes trouvés dans les préparations, on ignore combien d'entre eux étaient issus de la génération P. Ainsi, il se pourrait que dans les préparations témoins également, peu d'imagos soient demeurés indemnes par rapport à leur nombre initial, ce qui fait qu'il est impossible de procéder à une comparaison entre les résultats obtenus dans les échantillons comportant le prédateur et les valeurs témoins.
Outre la nature des animaux hôtes eux-mêmes, il semble également que le substrat nutritif qui leur est destiné constitue un facteur important pour le développement de T. nigrescens. C'est ainsi que l'on a pu observer dans 61 % des séries d'essais pratiquées sur du maïs (n=23; à l'exclusion toutefois des séries d'essais faisant intervenir P. truncatus) une reproduction de T. nigrescens (tabl. 4). Sur les substrats d'élevage habituels comme le son, le blé, les flocons d'avoine et le sorgho, on n'a trouvé en revanche de descendants de T. nigrescens que dans 38 % des essais (n=21; à l'exclusion des séries d'essais faisant intervenir P. truncatus).
S'agissant des cultures de O. mercator, T. nigrescens s'est uniquement reproduit lorsque ces animaux étaient élevés sur du mais, et non sur du son.
Sur les 10 préparations expérimentales impliquant R. dominica sur du sorgho, 4 seulement contenaient des descendants de T. nigrescens. Sur le maïs au contraire, T. nigrescens s'est reproduit efficacement dans les 10 échantillons contenant R. dominica comme animal hôte.
En comparaison du maïs, les imagos de R. dominica et de O. mercator ont pu se reproduire beaucoup mieux sur leur substrat d'élevage habituel. C'est ainsi qu'au cours de la première série d'essais avec R. dominica sur du sorgho, on a dénombré par ex. 7,5 fois d'adultes de cette espèce que dans les échantillons confectionnés parallèlement à partir de maïs. Le prédateur disposait par conséquent dans les échantillons de sorgho d'un nombre de proies très supérieur. En dépit de cela, les seuls descendants de T. nigrescens trouvés au cours de cette série d'essais l'ont été sur le maïs.
On peut en conclure que la capacité reproductive de T. nigrescens ne peut pas uniquement dépendre du nombre de proies disponibles, mais que le mais possède également en tant que milieu nutritif une fonction clé au regard de son développement.
R. dominica, S. zeamais, T. castaneum, pas plus que les deux souches de T. confusum n'ont pu se reproduire sur le manioc. P. truncatus, D. porcellus et O. surinamensis ne se sont reproduits sur les racines séchés que de façon négligeable.
L'hypothèse selon laquelle les racines de manioc provenant du Togo auraient pu contenir des résidus d'insecticides susceptibles d'avoir une action inhibitrice sur la capacité reproductive des coléoptères a été rendue improbable par une communication orale de HELBIG (1992). Selon ses informations, les racines de manioc séchées ne subissent en général pas de traitement aux insecticides.
T. stercorea s'est par ailleurs développé sur des racines de manioc provenant du même lot mieux que sur n'importe quel autre substrat. Nombreux étaient les imagos d'autres espèces de coléoptères testées qui, placés à l'origine sur les échantillons au manioc, étaient encore en vie 2 mois après le début de l'essai.
La plupart des espèces de coléoptères utilisées dans les expériences sur du manioc provenaient d'élevages de laboratoire et étaient nourries depuis de nombreuses années sur d'autres milieux nutritifs. Il est toutefois peu probable que les populations aient perdu en l'occurrence leur faculté première de se reproduire sur le manioc. Il semble donc que si le manioc fournit à la plupart des espèces de coléoptères ravageurs des stocks un bon substrat nutritif, il ne constitue pas un substrat d'incubation fiable pour les espèces de coléoptères considérées.
Là où il se trouvait en compagnie d'une autre espèce d'animal hôte, P. truncatus, a eu dans bien des cas des difficultés à se reproduire. C'est ainsi que dans les préparations comportant S. zeamais ou S. oryzae, par exemple, les espèces Sitophilus se sont nettement imposées face au bostryche. Il est certain que les charançons, qui, comme P. truncatus, font partie des ravageurs primaires, ont réussi en l'occurrence à concurrencer efficacement ce dernier dans la recherche de nourriture et l'occupation d'espaces d'incubation.
Tabl. 4: Capacité reproductive de T. nigrescens dans des cultures de coléoptères ravageurs des denrées stockées et influence du prédateur sur l'accroissement des populations
Le nombre moyen de descendants de T. nigrescens se situait entre:
*: 0,2 et 1,0 #: 5,4 et 10,0
**: 1,2 et 3,0 ##: 10,2 et 20,0
***: 3,2 et 5,8 ###: 20,2 et 36,8
- : pas de descendants de T. nigrescens
• : non effectué
/ : résultat du premier essai / second essai
Une action inhibitrice de T. nigrescens sur l'évolution des populations d'animaux hôtes était:
ND: non démontrable (p> 5,0 %)
PR : probable (p £ 5,0%)
SI : significative (p £ 1,0 %)
HS : hautement significative: (p £
0,1 %)
{ } : résultat pour les populations de P. truncatus
| Monocultures | Cultures
mixtes Sur mais avec {P.trun.} |
||||||||
| Animal hôte | Substrat d'élev. | Maïs | Manioc | ||||||
| D. porc. T | * | N | ** | N | ** | HS | # | HS | {HS} |
| R. domi. | -/** | N/N | ***/*** | N/N | * | FIS | ### | PR | {HS} |
| S. gr.af. | * | N | * | N | * | * | ### | N | {HS} |
| S. gr.gr. | * | N | * | N | * | * | ## | SI | {SI} |
| S. oryz. | *** | N | ** | N | * | * | ### | HS | {HS} |
| S. zeam. | * | * | * | N | * | N | ### | PR | {SI} |
| A. diap.T | * | * | * | N | * | * | ## | HS | {HS} |
| L. oryz. | - | N | - | N | * | * | ### | N | {HS} |
| L.oryz.T | * | SI | N | * | * | ### | HS | {HS} | |
| P. ratz. | - | N | * | N | * | * | ## | HS | {SI} |
| P. subd. | - | N | * | * | * | * | * | * | |
| T. cast. | - | N | - | N | - | N | # | HS | {N} |
| T. conf. | - | N | * | N | - | N | ## | HS | {SI} |
| T. conf. T | - | N | */- | PR/N | - | N | # | SI | {N} |
| A. adve. T | - | N | - | PR | * | * | ## | N | {HS} |
| C. pusi. | - | N | - | N | * | * | ### | HS | {SI} |
| O. merc. | - | HS | ** | HS | * | * | ## | HS | {HS} |
| O. surf. | *** | SI | *** | HS | * | SI | ## | N | {PR} |
| C. dimi. | - | PR | - | N | * | * | # | HS | {SI} |
| C. dimi.T | - | PR | - | N | * | * | # | N | {N} |
| T. gran. | - | SI | - | N | * | * | ### | PR | {HS} |
| T. ster. | ** | N | ** | N | ** | SI | # | N | {SI} |
Les séries d'essais impliquant S. granarius granarius montrent néanmoins que les espèces Sitophilus ne l'emportent pas dans tous les cas. Le rapport numérique des individus des deux populations était au cours de ces essais sensiblement équilibré. Dans les cultures mixtes comportant S. granarius africanus, c'est en effet P. truncatus, qui s'est imposé.
Dans les cultures mixtes, P. truncatus, a réussi à prendre l'avantage sur les deux autres espèces de Bostrychidés, à savoir D. porcellus et R. dominica.
En comparaison des résultats obtenus dans les monocultures sur du mais, les deux populations d'animaux hôtes se sont en général mal développées dans les cultures mixtes réunissant P. truncatus, et une des espèces de Ténébrionidés ou de Cucujidés, ainsi que l'une des souches de C. dimidiatus ou de T. stercorea.
Les deux essais réalisés par BÖYE (1988) montrent que les résultats des recherches entreprises sur des grains de mais en vrac ne peuvent pas être automatiquement appliqués au contexte des épis de mais stockés. Il a lui aussi mis en évidence le fait que sur les grains de mais en vrac, les populations de S. zeamais étaient avantagées par rapport à celles de P. truncatus, Si l'on place en revanche les deux espèces de coléoptères sur des épis de maïs, c'est la population de P. truncatus, qui connaît le développement le plus favorable.
Aussi bien dans les monocultures que dans les cultures mixtes comportant des ravageurs secondaires, on a ajouté dès le début des essais de la mouture de mais aux grains de mais servant de substrat. Considérant la présence simultanée de P. truncatus, en tant que ravageur primaire, il ne fallait pas s'attendre à un meilleur développement des ravageurs secondaires, ce qui n'a d'ailleurs pas été le cas.
Il semble que T. nigrescens ait davantage accepté comme proie la souche sauvage de L. oryzae provenant du Togo que la souche de laboratoire. En effet, alors que T. nigrescens n'a pu exercer d'influence sur l'évolution de la souche de laboratoire dans aucune des séries d'essais, le prédateur a, et cela aussi bien sur le son et dans la culture mixte, réduit significativement le nombre des imagos de L. oryzae qui faisaient partie de la souche sauvage provenant du Togo.
S'agissant des animaux hôtes de T. nigrescens, on n'a constaté aucune différence notable entre les souches de T. confusum et de C. dimidiatus élevées en laboratoire depuis de nombreuses années et les souches sauvages provenant du Togo.
Dans le cadre d'essais préliminaires, on a tenté de dégager une méthode adéquate de recherche sur la spécificité d'hôte de T. nigrescens (PÖSCHKO, 1992b). Les résultats variaient toutefois en fonction de l'approche choisie. Au contraire de résultats obtenus ultérieurement, on n'a pu observer dans les préparations expérimentales impliquant O. surinamensis ni de capacité de reproduction chez T. nigrescens, ni d'action inhibitrice du prédateur sur la population considérée.
Ceci explique que l'on ait choisi pour les recherches présentées ici une méthode unique, permettant de comparer directement les qualités des diverses espèces de coléoptères ravageurs des stocks susceptibles de servir d'animal hôte à T. nigrescens.
Les résultats parfois contradictoires obtenus malgré l'application de cette méthode unique soulignent bien la nécessité de procéder à de nombreux essais parallèles et de répéter des séries d'essais entières si l'on veut obtenir des données précises quant à la spécificité d'hôte de T. nigrescens. C'est ainsi par exemple que T. nigrescens. a pu se reproduire au cours d'une des séries d'essais mettant en oeuvre des cultures de R. dominica du sorgho, alors que la répétition n'a pas permis de trouver de descendants du prédateur dans les échantillons (tabl. 3. 1).
Il est certain que d'autres méthodes de recherche permettraient également de trouver des individus F1 isolés de T. nigrescens dans des élevages d'autres espèces de coléoptères qui n'ont pas été testées ici en tant qu'animaux hôtes, et vice-versa.
Si l'on tient par ailleurs compte de l'espérance de vie élevée des imagos de T. nigrescens et de la capacité de leurs femelles de pondre des oeufs plusieurs mois durant (3.2), il est probable que T. nigrescens serait en mesure de se multiplier en laboratoire sur d'autres animaux hôtes que P. truncatus,
Les résultats des recherches sur la spécifité d'hôte de T. nigrescens présentés ici ont été intégralement obtenus en laboratoire. On peut toutefois supposer que ces constatations ne sont applicables que sous certaines réserves aux conditions rencontrées sur le terrain.
Les animaux utilisés ont notamment été élevés dans des conditions climatiques constantes, favorables à leur développement. Même si les espèces d'insectes ravageurs des stocks vivant dans les greniers sont relativement bien protégées contre les variations climatiques saisonnières en raison du microclimat qui règne dans les épis de mais stockés en spathes, il n'en demeure pas moins qu'elles sont soumises à d'éventuels écarts de température, parfois extrêmes, comme il peut y avoir également des périodes de sécheresse ou d'humidité excessives, autant de conditions qui peuvent avoir des répercussions diverses sur l'évolution des coléoptères àl'intérieur des épis de mais.
Ni les prédateurs, ni les animaux hôtes n'avaient la possibilité de se soustraire au système imposé, pour se mettre par ex. à la recherche d'une source de nutrition plus désirable ou échapper à une surpopulation. Dans un grenier à maïs de construction traditionnelle, au contraire, les animaux peuvent en permanence émigrer ou faire place à d'autres espèces de ravageurs ou d'insectes utiles.
Les recherches en laboratoire continueront néanmoins de représenter une phase préliminaire indispensable, qui permet d'étudier, étape par étape, et d'appréhender les rapports complexes unissant le prédateur T. nigrescens àson hôte P. truncatus,
Venant s'ajouter à ceux de l'essai 3.3, les résultats des recherches exposées ici mettent en relief le rôle dominant de P. truncatus, en tant qu'animal hôte de T. nigrescens.
Au cours de la période couverte par les présentes recherches, T. nigrescens s'est reproduit uniquement dans les élevages de P. truncatus, placés sur du mais.
On a constaté en outre que T. nigrescens a pu se reproduire sur tous les substrats sur lesquels P. truncatus s'est développé.
T. nigrescens était également en mesure d'assurer sa reproduction là où les milieux choisis comme substrats d'incubation étaient moins favorables au développement de P. truncatus et où le nombre d'animaux hôtes disponibles était par conséquent moindre, comme par ex. sur le manioc et sur le blé.
T. nigrescens a inhibé de manière significative la croissance des populations de P. truncatus sur tous les substrats d'incubation testés sur le ravageur.
Dans les cultures mixtes associant dans chaque cas P. truncatus, à l'une des espèces de coléoptères sélectionnées, on a assisté en général à une forte multiplication de T. nigrescens. Outre qu'il a inhibé l'accroissement des populations de P. truncatus, T. nigrescens a également paralysé celle d'autres espèces de ravageurs.
Au vu des données actuelles, on peut raisonnablement penser que T. nigrescens va pouvoir se reproduire et se propager avec succès au Togo et en Afrique occidentale après qu'il aura été lâché.
Du fait que T. nigrescens n'a été jusqu'à présent observé dans la nature qu'en association avec P. truncatus, BÖYE 1988; REES, 1990), on va le retrouver uniquement dans les greniers à mais infestés par ce ravageur.
L'infestation provoquée par les ravageurs primaires attire d'autres espèces de ravageurs, comme les Ténébrionidés et les Cucujidés, qui ne sont pas par eux-mêmes en mesure d'endommager des grains de maïs intacts. On peut supposer que comme pour les essais de laboratoire, T. nigrescens va prendre pour proie principale les individus de P. truncatus, mais qu'il va également, dû à la présence de P. truncatus, détruire les oeufs et les larves d'autres espèces de ravageurs. Au milieu de ce brassage naturel d'espèces, composé de diverses espèces de coléoptères ravageurs des stocks, T. nigrescens va pouvoir se reproduire efficacement. Si la population du prédateur atteint une trop forte densité à l'intérieur du grenier, entraînant par là même au sein de cette population une certaine concurrence au niveau de la nourriture, ou si l'offre en nourriture se dégrade pour le prédateur, les T. nigrescens adultes vont alors émigrer et, s'aidant du phénomène des "allelochemicals", se mettre en quête d'un autre grenier à maïs infesté par leur animal hôte préféré, P. truncatus,
On peut donc partir du principe que la lutte biologique contre P. truncatus par la mise en oeuvre de T. nigrescens va être une réussite, qui va se traduire au bout du compte par une réduction des dégâts et pertes infligés aux stocks de maïs africains.
4.2.1
Introduction
4.2.2
Méthode
4.2.3 Résultats et commentaires
Outre quelque 60 espèces de coléoptères, on compte parmi les insectes ravageurs des denrées stockées environ une douzaine d'espèces de lépidoptères susceptibles de causer d'importants dommages économiques (WOHLGEMUTH, 1991)
Les recherches portant sur la spécificité d'hôte de T. nigrescens impliquaient d'une part les 4 principales espèces de pyrales: Ephestia cautella, E. elutella, E.. kuhniella et Plodia interpunctella, ainsi que le Galleriidé Corcyra cephalonica et le Géléchiidé Sitotroga cerealella (tabl. 2).
On a offert dans un premier temps en pâture à des imagos de T. nigrescens des oeufs de teignes dépourvus de substrat et placés dans de petites boîtes de Petri en verre.
Dans la deuxième phase de recherche, les oeufs de teignes ont été placés dans un bocal à essais, sur un substrat nutritif, ce qui a permis d'étudier l'influence de T. nigrescens sur l'évolution des populations de teignes.
Dans une troisième phase enfin, on a observé le développement d'une population mixte, composée d'oeufs de teignes de diverses espèces, avec présence simultanée de P. truncatus, et exposition ou non à l'action de T. nigrescens.
4.2.3 Résultats et commentaires
Les imagos de T. nigrescens ont dévoré les oeufs de teignes qui leur étaient proposés sans substrat.
Le prédateur n'a pas eu d'effets inhibiteurs sur l'évolution des populations de teignes implantées sur un substrat.
La présence simultanée de son animal hôte P. truncatus, n'a pas davantage permis de mettre en évidence une réduction du nombre de teignes de la part de T. nigrescens.
Si ces résultats démontrent que T. nigrescens est en mesure d'utiliser pour son alimentation les oeufs de certaines espèces de teignes ravageuses des stocks, il semble toutefois qu'il n'existe pas de mécanisme lui permettant de rechercher spécifiquement ce type de proie.
Considérant les résultats obtenus, on a décidé de renoncer à étudier plus avant la capacité de multiplication de T. nigrescens sur des lépidoptères comme animaux hôtes.
{kind=link}