3 Multiplication de Teretriosoma nigrescens avec Prostephanus truncatus comme animal hôte
Table
des matières - ![]() Précédente
- Suivante
Précédente
- Suivante![]()
3.1 Elevage de Prostephanus
truncatus et de Teretriosoma
nigrescens
3.2 Evolution démographique de Teretriosoma
nigrescens
3.3 Capacité reproductive de Teretriosoma
nigrescens dans des élevages de Prostephanus
truncatus sur divers substrats
3.4 Capacité reproductive de Teretriosoma
nigrescens dans des élevages de Prostephanus
truncatus sur un substrat végétal au bout
de plusieurs mois
3.1 Elevage de Prostephanus truncatus et de Teretriosoma nigrescens
3.1.1 Elevage de Prostephanus
truncatus
3.1.2 Elevage de Teretriosoma
nigrescens
On a testé dans le cadre d'essais préliminaires le taux de multiplication de T. nigrescens pour divers rapports numériques prédateur-proie. Il a été tenu compte en l'occurrence de l'âge des élevages de P. truncatus ainsi que de la composition connexe des différents stades d'évolution en tant qu'organismes destinés à servir de proies au prédateur. Les résultats de ces essais préliminaires nous ont conduit à élaborer les méthodes suivantes pour l'élevage de T. nigrescens et de P. truncatus
3.1.1 Elevage de Prostephanus truncatus
L'élevage de masse de l'animal hôte P. truncatus constituait la condition préalable à la multiplication du prédateur T. nigrescens. L'élevage de P. truncatus est pratiqué avec succès depuis 1983 au département de protection des récoltes de l'Institut fédéral de biologie de Berlin (BBA). Les populations d'origine avaient été importées à cet effet de Tanzanie. En partant de cette méthode d'élevage, on a placé env. 500 adultes de P. truncatus (ce qui correspond à peu près à un volume de 5 ml) sur 500 g env. de maïs en grains (provenant d'Argentine et commercialisé dans ce pays sous le nom de "La Plata"). Comme conteneurs d'élevage, on s'est servi de bocaux de verre de 2 litres de volume, après avoir soudé dans leur couvercle à fermeture par pression un morceau de treillis métallique (fig. 9A). Au bout de 4 à 5 semaines à 30°C ± 1°C et 75 % ± 5 % d'hum. rel., à l'abri de la lumière, on a assisté à l'éclosion de la génération F1 du ravageur. On disposait alors dans les bocaux d'élevage d'env. 2000 à 3000 P. truncatus adultes, lesquels ont été utilisés soit pour continuer l'élevage, soit pour des essais ou pour l'élevage de T. nigrescens.
3.1.2 Elevage de Teretriosoma nigrescens
La population d'origine de T. nigrescens a été importée en 1988 du Costa Rica. Les bocaux d'élevage, dans lesquels on avait placé des P. truncatus selon la méthode décrite précédemment, ont été soumis durant une semaine à une préincubation en chambre climatique, à l'abri de la lumière, là encore à 30°C ± 1°C et 75 % ± 5 % deum. rel. On a ensuite mis à pondre 50 imagos de T. nigrescens par bocal, ce qui correspond à un rapport prédateur-proie de 1 pour 10. Une semaine plus tard, on a retiré des élevages de P. truncatus les T. nigrescens de la première génération, de manière à pouvoir en disposer par la suite pour continuer l'élevage.
Huit semaines (à 30°C ± 1°C et 75 % ± 5 % d'hum. rel., à l'abri de la lumière) après avoir déposé les imagos de T. nigrescens sur les populations hôtes, on a pu retirer des bocaux d'élevage entre 70 et 150 imagos F1 de T. nigrescens, dont l'âge variait de 0 à 14 jours, et qui ont été utilisés pour des essais ou pour la continuation de l'élevage.
Le comptage des individus adultes F1 provenant de 28 bocaux d'élevage de T. nigrescens a permis d'établir une moyenne de 0,47 ± 0,15 descendants par femelle et par jour. S'appuyant sur les résultats exposés au chap. 3.2, on a admis ici que 50 % des imagos de T. nigrescens de la génération P étaient des femelles.
C'est ainsi que l'on a pu constituer dans un délai relativement court un élevage de masse de T. nigrescens. Les élevages de T. nigrescens, de même que les élevages connexes de P. truncatus, ont été aménagés de manière à ce que l'on puisse récupérer des coléoptères fraîchement éclos toutes les 2 semaines environ.
3.2 Evolution démographique de Teretriosoma nigrescens
3.2.1
Introduction
3.2.2 Matériels et méthodes
3.2.3
Résultats
3.2.4
Commentaires
Afin de déterminer le potentiel de reproduction d'une population de T. nigrescens, on a établi dans le cadre d'un essai de longue durée le nombre de descendants qu'une femelle de T. nigrescens pouvait produire au cours de sa vie.
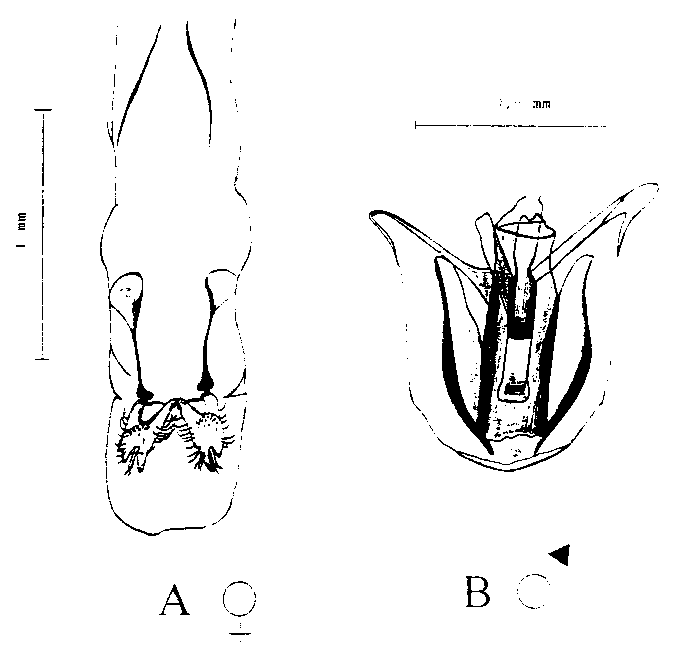
Pour pouvoir procéder à ces recherches, il fallait auparavant déterminer le sexe des individus de T. nigrescens. Etant donné que chez les imagos de T. nigrescens, il n'est pas possible de distinguer extérieurement les mâles des femelles, il a tout d'abord fallu confectionner une préparation pour pouvoir identifier les sexes au vu des parties génitales.
On a par ailleurs établi le rapport numérique entre les mâles et les femelles au sein d'une population de T. nigrescens.
Afin d'identifier l'appareil génital, on a tué un certain nombre de coléoptères adultes dans une solution à 70 % d'éthanol, puis on les a collés, sur la face ventrale, sur un porte-objet, ce qui a facilité la préparation des organes génitaux de ces animaux, qui sont extrêmement compacts. La préparation a été réalisée à un grossissement de 20 à 30 fois. Après le prélèvement, les organes génitaux ont été trempés durant quelques minutes dans une solution à 60 % env. d'acide lactique (C3H6O3), de manière à rendre les tissus environnants plus clairs. La comparaison avec les illustrations empruntées à TANNER (1925) a permis de déterminer lequel des deux appareils génitaux, qui diffèrent par leur forme, provenait d'une femelle.
Dans le but d'établir le rapport des sexes au sein d'une population de T. nigrescens, on a effectué 10 prélèvements successifs, en prenant à chaque fois dans les bocaux d'élevage 100 imagos de T. nigrescens, que l'on a tués dans une solution à 70 % d'éthanol. Des préparations écrasées, de souche, de ces animaux, ont ensuite été confectionnées. On a enfin placé sous un binoculaire les appareils génitaux mis à nu, afin de pouvoir distinguer les sexes.
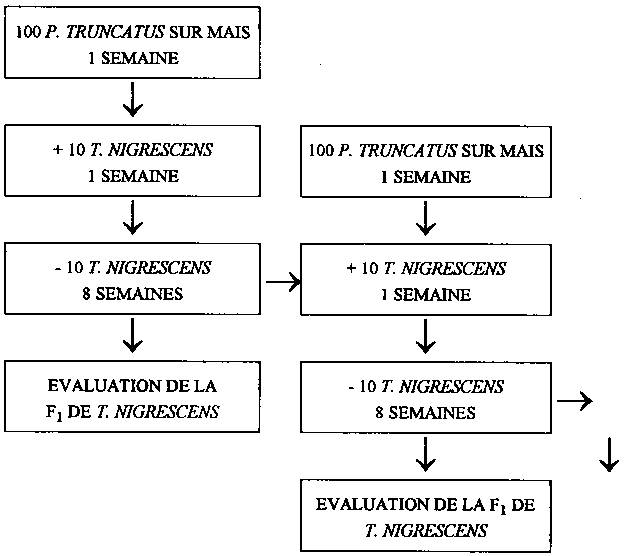
Dans le but de déterminer le nombre d'individus F1 qu'une femelle de T. nigrescens est capable d'engendrer, des populations de P. truncatus ont été sélectionnées en tant qu'élevages d'animaux hôtes et placées sur du maïs. 5 préparations parallèles ont été confectionnées, pour lesquelles on a introduit à chaque fois 100 imagos de P. truncatus dans des bocaux de verre à couvercle fileté, que l'on a placés sur 150 g environ de mais en grains. Ces préparations ont été mises à incuber en chambre climatique durant une semaine à 30°C ± 1°C et à 70 % ± 5 % d'hum. rel., à l'abri de la lumière, de manière à avoir des oeufs et de jeunes larves de l'animal hôte dans les échantillons au moment de l'introduction du prédateur. On a ensuite placé dans chaque bocal 10 imagos de T. nigrescens âgés de 0 à 2 semaines et de sexe indéterminé.
Les femelles de T. nigrescens ont eu sept jours durant la possibilité de pondre leurs oeufs sur ces élevages de P. truncatus. Les préparations ont été soumises pendant toute cette période aux conditions climatiques précédemment décrites. Les T. nigrescens adultes ont été ensuite éloignés des populations de l'animal hôte et placés de nouveau sur des élevages de P. truncatus datant de 7 jours.
En vue d'assurer le développement des oeufs de T. nigrescens, les populations d'animaux hôtes ont été placées en chambre climatique pour une nouvelle période de 8 semaines après que l'on retiré les imagos de T. nigrescens. On a ensuite dénombré les descendants de T. nigrescens (imagos et larves prénymphales) au sein des élevages de P. truncatus (fig 2).
Le rapport initial de 1 pour 10 entre les imagos du prédateur et ceux de sa proie a été maintenu constant pendant toute la durée des recherches. Lorsque l'un des 10 imagos de T. nigrescens de la génération P mourrait au cours d'une série d'essais, on limitait la population d'animaux hôtes suivante à 90 P. truncatus adultes. La quantité de maïs a été approximativement adaptée à l'importance de la population de P. truncatus. Cette quantité de mais n'a toutefois pas été réduite à moins de 50 g. et cela de façon à faciliter aux imagos de P. truncatus la pénétration dans les grains.
Suivant la méthode de préparation des organes génitaux décrite précédemment, les imagos de T. nigrescens de la génération P morts au cours des essais ont été examinés afin d'en déterminer le sexe.
Les imagos de T. nigrescens ont été constamment transférés sur de nouvelles populations de P. truncatus, jusqu'à ce que l'on ne trouve plus, lors de l'évaluation, de descendants du prédateur au sein des populations de P. truncatus pendant au moins 4 semaines consécutives.
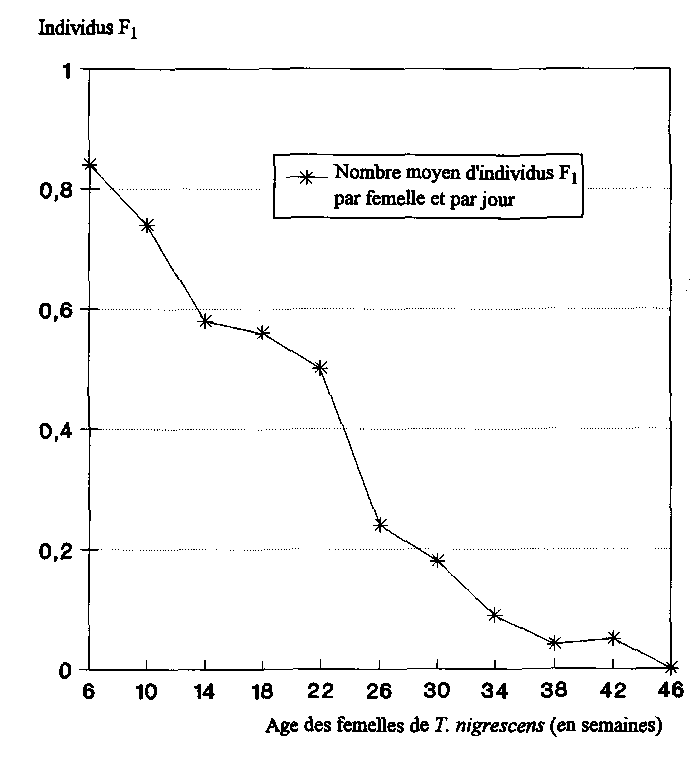
A la fin des essais, on a procédé à la détermination du sexe de tous les imagos de T. nigrescens encore en vie. Après avoir établi le nombre de femelles présentes dans chaque série d'essais, on a calculé le nombre moyen des descendants produits quotidiennement par une femelle de T. nigrescens. Pour ce faire, on a calculé tout d'abord les valeurs moyennes établies à partir de quatre observations consécutives pour chacune des séries d'essais, ce qui correspondait à chaque fois à une période d'essai de 4 semaines (28 jours). De manière à obtenir une synthèse des résultats des 5 séries d'essais, on a établi ensuite la moyenne générale de ces valeurs moyennes, ce qui fait que chacune des données indiquées à la fig. 4 a été calculée à partir de 20 valeurs partielles.
La figure 3 représente les organes génitaux respectifs du mâle et de la femelle de T. nigrescens. La préparation des parties génitales de 1000 imagos de T. nigrescens a révélé que le rapport entre les sexes était pratiquement de 1: 1 (52,0 ± 2,5 mâles contre 48,0 ± 2,5 femelles).
Dans 4 des 5 séries d'essais à long terme, on a dénombré à chaque fois 4 femelles de T. nigrescens, tandis que l'on a trouvé dans la cinquième série 5 femelles du prédateur. Parmi les 50 imagos de T. nigrescens de la génération P utilisés à l'origine, 21 étaient par conséquent des femelles.
Le nombre de descendants produits journellement par les femelles baissait au fur et à mesure qu'elles avançaient en âge (fig. 4). C'est avec des femelles de T. nigrescens âgées de 6 semaine au plus que l'on a déterminé la valeur moyenne maximale de 0,84 ± 0,19 individus Fi par femelle et par jour.
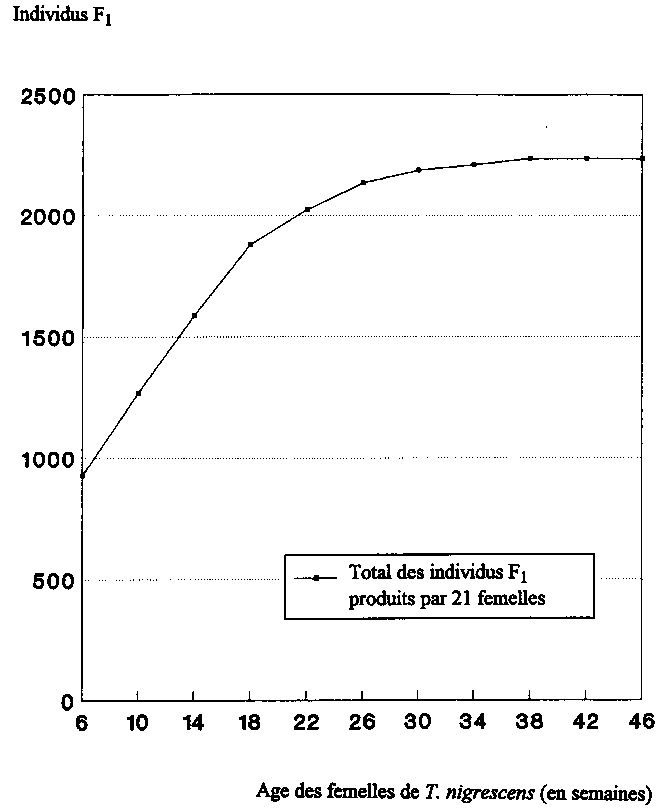
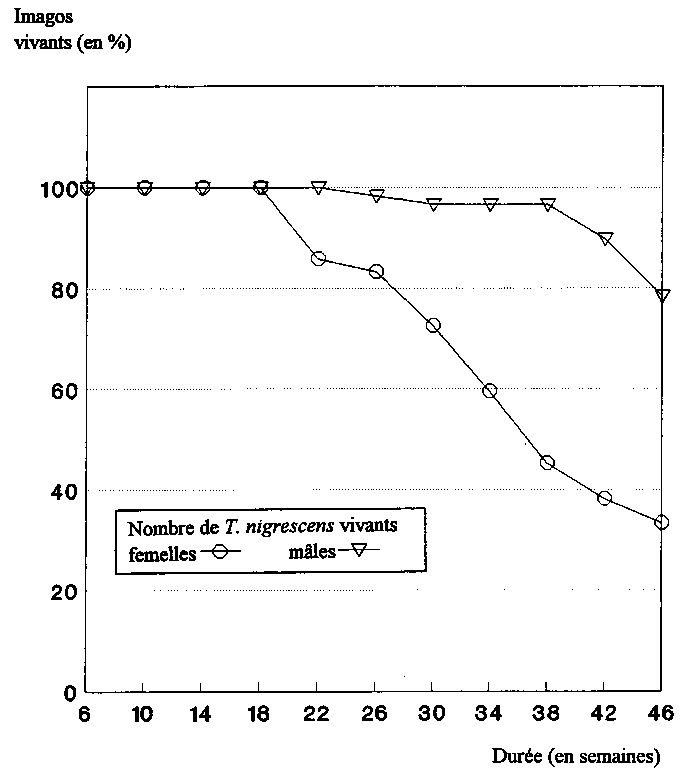
Les premières femelles de T. nigrescens sont mortes au bout de 18 semaines. Au sein des populations de P. truncatus, le nombre de descendants avait baissé jusqu'alors de 33 %, s'établissant à 0,56 ± 0,08 individus F1 par femelle et par jour. Au bout de 46 semaines, on n'a plus trouvé aucun descendant des 7 femelles de T. nigrescens encore vivantes (fig. 5).
Une femelle de T. nigrescens était donc en mesure de produire en moyenne 107 descendants au cours de sa phase reproductive.
Dans les conditions données, les femelles de T. nigrescens sont mortes plus tôt que les mâles (fig. 6). Durant les 38 premières semaines, il n'y a eu qu'un seul mort parmi les 29 mâles. A la fin des essais, 78,4 % des mâles et 33,3 % des femelles étaient encore en vie. Au bout de 46 semaines (10,7 mois), 59,5 % des 50 imagos de T. nigrescens utilisés étaient encore en vie, les deux sexes confondus.
Fig. 6: Espérance de vie des imagos de T. nigrescens sur des populations de P. truncatus
Dans le cadre de cet essai, on a saisi le nombre des individus F1 provenant de la ponte de T. nigrescens et ayant réussi à évoluer respectivement jusqu'au stade de larve prénymphale ou jusqu'à l'imago. Les oeufs pondus ne donnent pas forcément tous par la suite des animaux adultes. Il existe à l'intérieur des cycles évolutifs des individus d'une même population des taux de mortalité naturels. On peut par conséquent supposer que les femelles de T. nigrescens avaient pondu un nombre d'oeufs supérieur à celui des individus F1 observés.
La comparaison de données suivante révèle que l'âge "absolu" des femelles de T. nigrescens n'est pas le seul facteur déterminant en ce qui concerne le nombre de descendants produits. Les conditions dans lesquelles ont été élevés les animaux avant et pendant la ponte exercent en effet une certaine influence sur l'âge "relatif" des femelles.
Dans les préparations entrant dans le cadre des essais pratiqués avec T. nigrescens sur un substrat végétal (cf. 5.2.1), les imagos ont été transférés à intervalles de 4 semaines dans de nouveaux élevages de P. truncatus. Chez les femelles de T. nigrescens, on a pu constater au bout de 15 mois encore la présence de descendants.
Dans les cas où les imagos de T. nigrescens ont été placés sur du mais en l'absence d'animaux hôtes (cf. 3.4), certaines femelles de T. nigrescens âgées de 16,5 mois étaient encore en mesure de produire des descendants après leur transfert sur des populations de P. truncatus, alors qu'à l'inverse, pendant les essais que nous venons de décrire, des femelles de T. nigrescens âgées de 10,7 mois ne produisaient déjà plus de descendants.
Il est probable que ces femelles ont trouvé dans l'offre sans cesse renouvelée en cultures de P. truncatus des conditions de ponte optimales. Comparé aux animaux n'ayant produit aucune descendance, l'énorme dépense d'énergie nécessaire à la production des oeufs s'est traduit sur le plan physiologique par un vieillissement accéléré des femelles. Ce phénomène pourrait également expliquer la mort relativement rapide des femelles de T. nigrescens par rapport aux mâles, ainsi que la haute longévité de ces animaux sur des substrats végétaux.
Il est peu probable que les résultats différents obtenus au cours des recherches sur la capacité reproductive et la longévité de T. nigrescens soient à rattacher à des fluctuations naturelles, spécifiques à certains individus au sein d'une population. Les résultats montrent que lors des recherches sur les taux de ponte, il est nécessaire d'une part de connaître l'âge des femelles utilisées, et d'autre part de tenir compte des conditions d'élevage des animaux.
3.3.1
Introduction
3.3.2 Matériels et méthodes
3.3.3
Résultats
3.3.4
Commentaires
Les recherches effectuées jusqu'à présent sur la capacité de prédation et de reproduction de T. nigrescens sur P. truncatus en tant qu'animal hôte ont été menées sur du maïs. La plupart des ravageurs des stocks ne sont cependant pas dépendants d'un substrat d'incubation déterminé. Outre le maïs, P. truncatus est lui aussi capable de se reproduire sur d'autres substrats. BÖYE (1990) a pu mettre par exemple en évidence une multiplication de P. truncatus sur le manioc, le sorgho et le riz.
Les expériences suivantes avaient pour but de vérifier si T. nigrescens était également en mesure d'inhiber l'accroissement d'une population de P. truncatus sur d'autres substrats que le maîs et s'il pouvait s'y reproduire.
On a utilisé comme substrats du manioc, du blé, du sorgho, des pois et du son (le son de blé additionné de glycérine, de glucose et de levure s'est avéré un sustrat d'élevage tout à fait approprié pour de nombreuses espèces de coléoptères et de lépidoptères ravageurs des stocks).
Pour chaque série d'essais, on a rempli dix bocaux de verre de 300 ml jusqu'à la moitié environ, en y introduisant 150 g de blé, de sorgho ou de pois, ou encore 100 g de manioc ou 50 g de son de blé. On ensuite placé dans chaque bocal 100 imagos de P. truncatus. A l'issue d'un temps d'incubation de 7 jours en chambre climatique, à 27° ± 1°C et 75 % ± 5 % d'hum. rel., à l'abri de la lumière, on a introduit dans la moitié des bocaux de chaque série d'essais (n=5) 10 jeunes imagos de T. nigrescens. Les autres préparations, qui ne contenaient donc pas de prédateurs, servaient de témoins. L'évaluation est intervenue huit semaines après l'introduction des T. nigrescens adultes. On a dénombré en l'occurrence dans les échantillons les imagos de P. truncatus et les descendants de T. nigrescens. Les valeurs de comparaison ont été fournies par les résultats de deux autres séries d'essais effectués avec P. truncatus sur du mais. P. truncatus étant décrit également comme ravageur du manioc, on a procédé à deux séries d'essais en utilisant des racines de manioc provenant du Togo.
L'interprétation statistique des résultats d'essais a été réalisée à l'aide du test t, selon RENNER (1981). En partant du nombre d'imagos de l'animal hôte dans chacun des 5 échantillons, sans prédateurs et avec prédateurs, on a calculé dans un premier temps la moyenne arithmétique (m.a.), en tenant compte de l'écart type (e.t.). La comparaison de ces valeurs moyennes (test t) a ensuite permis d'établir si T. nigrescens exerçait ou non une influence sur l'évolution de la population de coléoptères.
Lorsque les valeurs moyennes divergaient avec une probabilité d'erreur
| • de > 5 % | l'influence de T. nigrescens a été qualifiée de non démontrable |
| • de £ 5 % | l'influence de T. nigrescens a été qualifiée de probable |
| • de £ 51 % | l'influence de T. nigrescens a été qualifiée de significative |
| • de £ 0,1 % | l'influence de T. nigrescens a été qualifiée de hautement significative. |
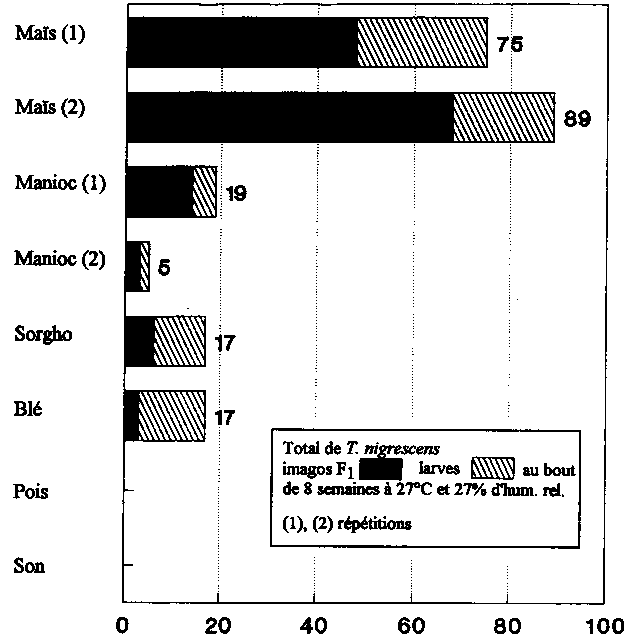
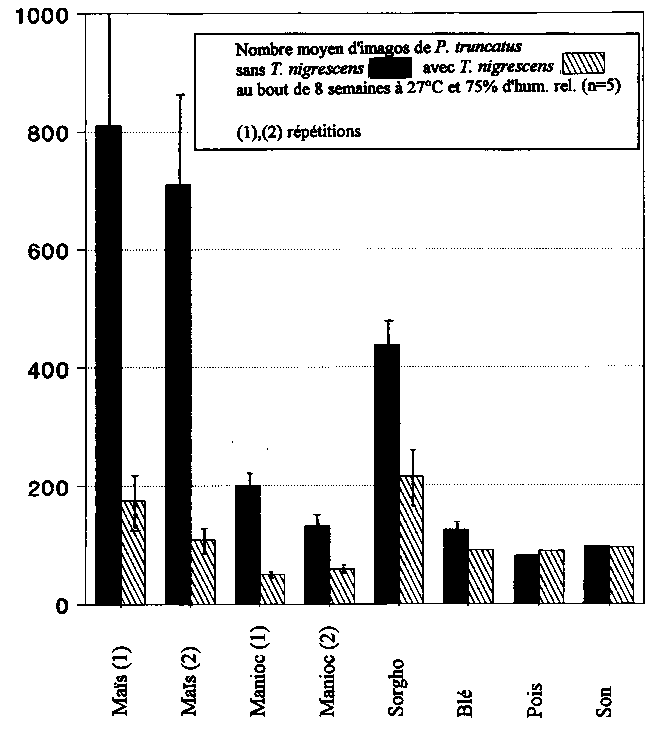
C'est dans les préparations comparées utilisant du maïs que P. truncatus a trouvé les meilleures conditions de reproduction (tabl. 1). Les populations d'animaux hôtes se sont multipliées jusqu'à atteindre 7 à 8 fois leur nombre initial de 100 imagos. Dans les échantillons contenant T. nigrescens, on a trouvé dans la première et dans la seconde série d'essais respectivement 78 et 85 % d'imagos de P. truncatus de moins que dans les préparations témoins sans prédateurs, deux chiffres hautement significatifs. 48 imagos et 27 larves de T. nigrescens ont été dénombrés au cours du premier essai, tandis que l'on a compté au total lors de la répétition 61 imagos F1 et 21 larves.
P. truncatus n'a pu se reproduire sur le manioc que de façon médiocre. Pour l'une des séries d'essais, on a dénombré en moyenne environ 200 imagos par préparation, et de 110 à 150 dans l'autre (tabl. 1). T. nigrescens a permis d'arriver à une réduction très significative de 75 et 55 % du nombre d'adultes de l'animal hôte (fig. 7). T. nigrescens n'a pratiquement pas pu se reproduire dans ces élevages de P. truncatus On n'a en effet dénombré ici que 19 descendants du prédateur.
P. truncatus s'est relativement bien reproduit sur le sorgho (tabl. 1). On a dénombré entre 380 et 450 imagos de P. truncatus par préparation expérimentale. Par rapport aux témoins, T. nigrescens a réduit très significativement le nombre des adultes de l'animal hôte de 51 % (fig. 7). Au total, on a trouvé dans ces élevages 6 imagos F1 et 11 larves de T. nigrescens.
Le blé s'est revélé pour P. truncatus un substrat d'incubation médiocre. Dans les élevages sans prédateurs, on a dénombré seulement de 110 à 150 imagos par préparation (tabl.), tandis que l'on comptait dans les élevages avec Y nigrescens une réduction significative de 27 % du nombre d'adultes de P. truncatus (fig. 7). On a trouvé au total 3 imagos F1 et 14 larves du prédateur.
Sur les pois et le son, enfin, il n'y a pas eu multiplication de P. truncatus et aucun descendant de T. nigrescens n'a été trouvé dans les échantillons. Il n'a pas été possible de constater de la part de T. nigrescens des effets réducteurs sur le nombre des adultes de l'animal hôte (tabl. 7). Sur les pois, on a même trouvé dans les échantillons avec T. nigrescens significativement plus d'imagos de P. truncatus que dans les préparations témoins.
Tabl. 1: Influence et capacité reproductive de 10 T. nigrescens au sein de populations de P. truncatus
100 imagos sur divers substrats; évaluation au bout de 8 semaines a 27°C et 75 % d'hum. rel. (m.a. ± e.t., n=5)
*: tous les P. truncatus étaient morts; (1), (2): répétitions; SI, HS: différence significative (p£ 1%), hautement significative (p<0,1%) avec témoin sans T. nigrescens.
| Préparation | Adultes P. trunc. |
Descendance T. Nigr. |
Substrat | ||
| P.trun. | (1) | 810,2 ± 190,6 | HS | Maïs | |
| P.trun. + T.nigr. | 175,6 ± 52,3 | 15,0 ± 4,9 | |||
| P.trun. | (2) | 710,8 ± 153,2 | HS | Maïs | |
| P.trun. + T.nigr. | 109,6 ± 25,1 | 17,8 ± 2,2 | |||
| P.trun. | (1) | 200,4 ± 190 | HS | Manioc | |
| P.trun. +T.nigr. | 50,4 ± 7,8 | 3,8 ± 2,0 | |||
| P.trun. | (2) | 132,6 ± 17,4 | HS | Manioc | |
| P.trun. + T.nigr. | 59,4 ± 12,0 | 1,0 ± 0,7 | |||
| P.trun. | 438,0 ± 40,0 | HS | Sorgho | ||
| P.trun. + T.nigr. | 215,8 ± 50,6 | 3,4 ± 2,2 | |||
| P.trun. | 125,2 ± 16,0 | SI | Blé | ||
| P.trun. + T.nigr. | 91,2 ± 8,4 | 3,4 ± 2,8 | |||
| P.trun. | 81,6 ± 2,6* | SI | Pois | ||
| P.trun. + T.nigr. | 90,6 ± 4,8* | 0 | |||
| P.trun. | 97,6 ± 1,8 | Son | |||
| P.trun. + T.nigr. | 96,2 ± 1,3 | 0 | |||
Fig 8: Influence de T. nigrescens sur l'évolution de populations de P truncatus sur divers substrats
Des descendants de T. nigrescens ont été trouvés sur tous les substrats sur lesquels P. truncatus s'est reproduit (mais, manioc, sorgho et blé).
Toutefois, le prédateur n'a pu se multiplier (nombre d'animaux de la génération P/nombre d'individus F1 > 1) au cours de la période d'essai de 8 semaines que dans les cas où les populations d'animaux hôtes étaient élevées sur du mais. Il est probable que l'abondance de la nourriture offerte est à l'origine du fait que le prédateur a pu se multiplier davantage qu'au sein des autres populations d'animaux hôtes.
T. nigrescens a pu se reproduire également sur les élevages de P. truncatus lorsque le milieu utilisé comme substrat d'incubation était moins bien adapté à P. truncatus ce qui est par ex. le cas pour le manioc et le blé, en dépit de la réduction du nombre de proies que cela impliquait. T. nigrescens était en mesure d'inhiber l'accroissement des populations de P. truncatus sur tous les substrats d'incubation testés sur le ravageur. L'efficacité de T. nigrescens dans la lutte contre P. truncatus n'est donc pas seulement limitée en l'occurrence au mais, mais se retrouve également sur d'autres substrats victimes du ravageur.
3.4.1
Introduction
3.4.2 Matériels et méthodes
3.4.3
Résultats
3.4.4
Commentaires
On n'a pas observé de reproduction de T. nigrescens sur des substrats végétaux en l'absence d'animaux hôtes (5.4). C'est dans le but de vérifier s'il s'agissait d'une pause dans la reproduction ou au contraire d'une perte irréversible de la capacité reproductive que l'on a procédé aux recherches décrites dans la suite.
A l'issue d'une période de 9,5 mois, on a retiré de chacune des préparations expérimentales décrites au chap. 5.2 (n=11), dans lesquelles vivaient encore de nombreux T. nigrescens (mais, manioc, haricots, sorgho, blé, flocons d'avoine, cacahuètes, amandes et riz), 10 coléoptères (tabl. 8), que l'on a placés dans des élevages de P. truncatus commencés 7 jours auparavant (100 P. truncatus adultes sur env. 150 g de mais, dans des bocaux de verre de 300 ml). En tant que témoin, on a collecté de jeunes T. nigrescens dans des bocaux d'élevage, lesquels ont été eux aussi placés sur de nouveaux élevages d'animaux hôtes (n=5). Des élevages de P. truncatus ne contenant pas de prédateurs servaient de témoin supplémentaire (n=5). A l'issue d'un temps d'incubation de 8 semaines en chambre climatique à 27° ± 1°C et 75 % ± 5 % d'hum. rel., à l'abri de la lumière, on a procédé au dénombrement de la descendance de T. nigrescens et des P. truncatus adultes dans les préparations expérimentales.
Un second essai a eu lieu au bout de 16 mois. Les coléoptères avaient été en l'occurrence prélevés sur des échantillons comportant du maïs, du manioc, du blé et des flocons d'avoine (n=6). L'évaluation est intervenue après 12 semaines.
A l'issue d'une pause de 9,5 mois, T. nigrescens était à nouveau en mesure de se reproduire sur des populations de son animal hôte, P. truncatus Dans les bocaux à essai, on comptait en moyenne au bout de 2 mois 9,5 5,0 descendants de T. nigrescens (imagos et larves). Corollairement, les coléoptères avaient conservé leur capacité de réduire l'accroissement des populations d'animaux hôtes. Dans les témoins sans T. nigrescens, on a dénombré en moyenne 810,2 ± 190,6 adultes de P. truncatus alors que les bocaux dans lesquels le ravageur était présent ne contenaient que 250,7 ± 90,4 animaux hôtes, ce qui correspond à une réduction de 69 %. Avec 15,0 ± 4,9, le nombre moyen de descendants de T. nigrescens trouvés dans les échantillons renfermant de jeunes animaux témoins provenant des bocaux d'élevage était légèrement plus élevé. Dans ces préparations expérimentales, on ne comptait plus que 175,6 ± 52,3 adultes de P. truncatus Comparé aux témoins sans T. nigrescens, cela représente une réduction de 78 %.
Même au bout de 16 mois d'existence en milieu nutritif végétal, T. nigrescens était encore capable de se reproduire sur des élevages de P. truncatus et d'inhiber l'accroissement des populations du ravageur. Après 3 mois, les échantillons contenaient en moyenne 30,7 ± 12,2 descendants de T. nigrescens et 158,8 ± 74,5 P. truncatus adultes. Les populations de P. truncatus qui avaient pu évoluer librement avaient atteint en moyenne 653,3 ± 193,6 imagos, ce qui correspond à une réduction de 76 % environ sous l'action de T. nigrescens.
Ces résultats s'expliquent par un phénomène de cannibalisme. Dû à la pénurie de nourriture, les imagos de T. nigrescens dévorent leurs propres oeufs. Ce n'est qu'en présence d'un nombre de proies suffisantes qu'ils cessent de manger leurs oeufs (voir aussi à ce sujet 5.2).
Il existe une autre thèse, selon laquelle une alimentation purement végétale déclencherait chez T. nigrescens une pause réversible dans la reproduction. La carence en protéines animales, cholestérol ou autres substances nutritives essentielles absentes d'une alimentation végétale ou ne s'y trouvant qu'en quantités insuffisantes pourrait entraîner une inhibition de l'oogénèse. Dès que la substance nutritive recommence d'être absorbée à travers l'alimentation animale, le processus de maturation des oeufs se remet également à fonctionner.
On peut supposer que les femelles de T. nigrescens s'étaient déjà accouplées au moment de leur introduction dans les bocaux d'essais contenant les substrats végétaux. Traversant l'orifice génital de la femelle, le sperme du mâle pénètre dans le receptaculum seminis, qui est une sorte de réservoir à semence Les oeufs parvenus à maturité glissent alors le long du réceptacle, pour être ensuite fécondés dans l'ovipositeur de la femelle (WEBER & WEIDNER, 1974). Ainsi, il suffit souvent d'un seul accouplement pour assurer la fécondation de tous les oeufs en cours de maturation d'une femelle. L'inhibition de la spermatogénèse sur un substrat végétal ne joue par conséquent qu'un rôle assez secondaire.
Les prédateurs sont très rapidement capables de pondre de nouveau des oeufs fécondés. C'est ainsi que des imagos de 71 nigrescens placés durant 3 mois sur une mouture de mais étaient déjà prêts en l'espace de 7 jours à déposer de nouveau des oeufs sur des élevages de P. truncatus permettant ainsi le développement ultérieur de T. nigrescens adultes.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}