![]()
![]()
![]()
caractéristiques des fruits à la récolte
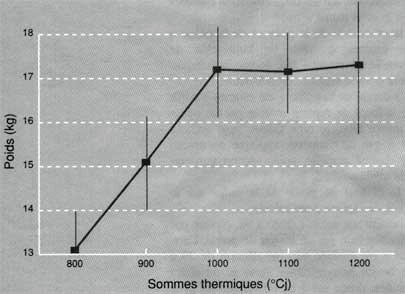
Le poids moyen des régimes a augmenté de façon très importante entre les stades 800 et 1 000 °Cj de somme thermique, puis il s'est plus ou moins stabilisé (figure 1). Après 1000 °Cj, le ralentissement observé serait causé, en grande partie, par une diminution de l'activité photosynthétique du au vieillissement des feuilles, d'une part, et à leur nombre de moins en moins élevé, d'autre part (tableau I).
Après les stades 900 et 1000 °Cj, la dureté de la pulpe diminue (figure 2) et le rapport du poids de la pulpe sur le poids de la peau augmente plus rapidement (figure 3). Les taux de matière sèche dans les fruits croissent jusqu'au stade 1000 °Cj; pour les récoltes de plus en plus tardives les teneurs baissent (figure 4).
La coloration de la pulpe s'accentue au fur et à mesure que le stade de coupe est retardé; la couleur est le plus souvent plus prononcée dans les deuxièmes mains que dans les quatrièmes (figure 5), traduisant un développement plus avancé des premières mains en général; ces observations sont en accord avec les résultats obtenus sur d'autres cultivars de bananes plantains (COLLIN, 1991; Cirad-Irfa, 1991).
L'extrait sec soluble et l'acidité - de 5,5 % et 4,8 meq/100 g de matière fraîche, respectivement - restent constants jusqu'au stade 1100 °Cj. À 1200 °Cj, l'acidité augmente légèrement (5,1 meq/100 g de matière fraîche), ce qui caractérise l'entrée des fruits en phase de mûrissement.
conservation des fruits
À température ambiante, il faut environ 18 à 23 j pour que 50 % des lots récoltés au stade de maturité de 800 à 900 °Cj parviennent à maturation; pour les récoltes plus tardives à 1 200 °Cj, 5 à 6 j suffisent pour obtenir le même résultat (figure 6). À température ambiante, la durée de conservation des fruits correspondant à l'observation de 100 % de doigts mûrs est sensiblement moitié moindre que celle enregistrée dans le cas d'un stockage au froid (figure 7). Ce résultat serait en accord avec la règle du Q10 selon laquelle la vitesse d'une réaction chimique ou enzymatique serait doublée lorsque la température du milieu serait augmentée de 10 °C (HELLER, 1969); dans l'expérimentation présentée, les températures de conservation testées, 24-27 °C et 12 °C, sont effectivement proches des conditions requises pour vérifier la règle.
Pour des séjours au froid prolongés au-dessus de 30 à 35 j la coloration de la peau a montré une évolution anormale au fur et à mesure de la maturation des doigts qui présentaient, alors, fréquemment, des plages sombres, semblables à celles constatées par NGALANI (1986). À conditions de conservation identiques, les taux d'extraits secs solubles et les acidités ont été semblables quels que soient le doigt et le stade de récolte considérés.
Pour une conservation à température ambiante, à stades identiques de coloration de la peau, le taux d'extrait sec soluble (31 %) et l'acidité des pulpes (16 meq/100 g de matière fraîche) des plantains mûrs ont été supérieurs à ceux des fruits stockés au froid, qui ont donné, respectivement, 28 % et 10 meq/100 g de matière fraîche.
Les pertes de poids sont plus restreintes au froid (moins de 1 % et 3 à 4 % après 10 et 20 j) qu'à température ambiante (6 à 8 % puis 15 %) quand les doigts sont encore verts; le mûrissement accentue la diminution de poids (figure 8).
![]()
![]()
![]()
{kind=link}
{kind=link}